


Czym jest teoria hologenomu?
Rosyjski botanik i teoria hologenomu
|
|

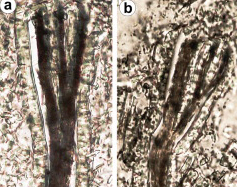
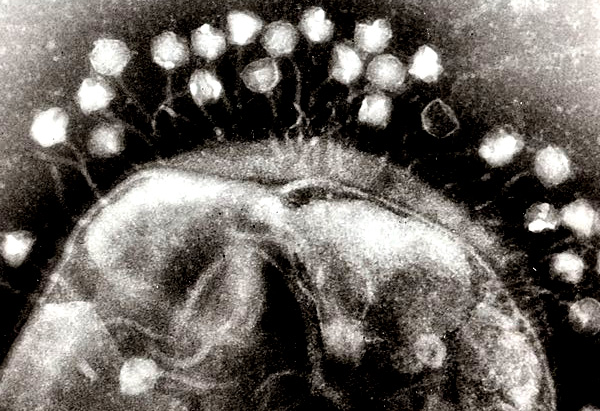
Pojęcie holobiontu (ze starożytnej greki: hólos – cały, biont – jednostka życia) zostało wprowadzone w latach 90. XX wieku przez Lynn Margulis i oznacza zespół organizmów żyjących w symbiozie przez istotną część ich historii życiowej, tworzących ekologiczną strukturę i mogących podlegać selekcji naturalnej [15]. Ostatnie twierdzenie rozwijane jest w ramach teorii hologenomu. Mówi ona, że genom gospodarza (np. człowieka) i mikroorganizmów zamieszkujących dostępne w nim nisze ekologiczne (np. przewód pokarmowy) stanowić mogą wyższej kategorii jednostkę fizjologiczno-ekologiczno-ewolucyjną. Z punktu widzenia gospodarza jego mikrobiom jelitowy uznać można za swego rodzaju kolejny narząd, spełniający fizjologiczno-regulacyjne funkcje, natomiast gospodarz jest dla mikroorganizmów siedliskiem [16].
|
|

Mikrobiom a hologenom
Więzi między wielokomórkowcami w teorii hologenomu

nowe znaczenie mikrobiomu w medycynie.








W czasie czytania tego artykułu nasunęło mi się takie pytanie: Czy w takim razie za holobionty można uznać np. porosty?
Marcin Piotr
Jejku! Uwielbiam takie artykuły na Twoim blogu 😉
Male sprostowanko: „Dla przeżuwaczy niezwykle ważny jest mikrobiom składający się z bakterii, archeanów, grzybów i pierwotniaków, rozkładający w żwaczu celulozę do kwasów tłuszczowych, białek i gazów” – to zdanie mija sie z prawda, celuloza jako taka to polimer cukrowy. Ani w niej bialek ani tluszczy. Ktores z wyrazen do poprawy 🙂
Bardzo ciekawy artykul, dzieki! Faktycznie ciezko chyba bedzie zaprzeczyc, ze skoro organizm wielokokorkowy i jego mikrobiom sa raczej nierozerwalne, to dobor naturalny dziala na cala jednostke rownoczesnie (choc na pewno wsrod mikrobiomu odbywa sie osobny dobor w zamknietym srodoeisku hosta).
Male sprostowanko: „Dla przeżuwaczy niezwykle ważny jest mikrobiom składający się z bakterii, archeanów, grzybów i pierwotniaków, rozkładający w żwaczu celulozę do kwasów tłuszczowych, białek i gazów” – to zdanie mija sie z prawda, celuloza jako taka to polimer cukrowy. Ani w niej bialek ani tluszczy. Ktores z wyrazen do poprawy 🙂
Bardzo ciekawy artykul, dzieki!
Dzięki, złego sformułowania użyłem. Poprawione.